Сенсоры различают по специфичной чувствительности к различным стимулам, по строению и местоположению. Специфичная чувствительность рецепторов к раздражителям различной природы (механическим, хим, температурным и т. д.) обоснована различными механизмами управления ионными каналами плазматических мембран, состояние которых определяет появление рецепторного потенциала и переход от физиологического покоя к возбуждению. Стимулы, к которым сенсоры более чувствительны, именуются адекватными (лат. adaequatus — приравненный).
Механорецепторы возбуждаются посильнее всего вследствие деформации их клеточной мембраны при давлении либо растяжении, к ним относятся тактильные сенсоры кожи, проприоцепторы мускул и сухожилий, слуховые и вестибулярные сенсоры во внутреннем ухе, барорецепторы и волюморецепторы, находящиеся во внутренних органах и кровяных сосудах. Хеморецепторы возбуждаются вследствие присоединения к ним определенных хим молекул, они представлены обонятельными и вкусовыми сенсорами, также хемочувствительнымирецепторами внутренних органов и кровеносных сосудов.
Для расположенных в сетчатке глаза фоторецепторов адекватным раздражителем являются поглощенные ими кванты света, для терморецепторов (холодовых и термических) — конфигурации температуры.
В итоге действия раздражителя практически у всех рецепторов возрастает проницаемость клеточной мембраны для катионов, что приводит к ее деполяризации. Исключением из общепринятого правила являются фоторецепторы, где после поглощения энергии квантов света в связи особенностями управления ионными каналами происходит гиперполяризация мембраны. Изменение величины мембранного потенциала рецепторов в ответ на действие стимула представляет собой рецепторный потенциал — входной сигнал первичных сенсорных нейронов. Если величина рецепторного потенциала достигнет критичного уровня деполяризации либо превзойдет его, генерируются потенциалы деяния, при помощи которых сенсорные нейроны передают в центральную нервную систему информацию о действующих стимулах.
Информация о силе действующего на сенсоры стимула кодируется 2-мя методами: частотой потенциалов деяния, возникающих в сенсорном нейроне (частотное кодирование), и числом сенсорных нейронов, возбудившихся в ответ на действие стимула. При увеличении силы действующего на сенсоры раздражителя увеличивается амплитуда рецепторного потенциала, что, обычно, сопровождается повышением частоты потенциалов деяния в сенсорном нейроне первого порядка. Чем обширнее имеющийся частотный спектр потенциалов деяния у сенсорных нейронов, тем большее число промежных значений силы раздражителя способна различать сенсорная система. Первичные сенсорные нейроны схожей модальности различаются порогом возбуждения, потому при действии слабеньких стимулов возбуждаются только более чувствительные нейроны, но с повышением силы раздражителя на него реагируют и наименее чувствительные нейроны, имеющие более высочайший порог раздражения. Чем больше первичных сенсорных нейронов возбудится сразу, тем посильнее будет их совместное действие на общий нейрон второго порядка, что в конечном итоге отразится на личной оценке интенсивности действующего раздражителя.
Продолжительность чувства находится в зависимости от реального времени меж началом и прекращением воздействия на сенсоры, также от их возможности уменьшать либо даже прекращать генерацию нервных импульсов при длительном действии адекватного стимула. При продолжительном действии стимула порог чувствительности рецепторов к нему может повышаться, что определяется как адаптация рецепторов. Механизмы адаптации не схожи в сенсорах различных модальностей, посреди их различают стремительно адаптирующиеся (к примеру, тактильные сенсоры кожи) и медлительно адаптирующиеся сенсоры (к примеру, проприоцепторы мускул и сухожилий). Стремительно адаптирующиеся сенсоры посильнее возбуждаются в ответ на резвое нарастание интенсивности стимула (фазический ответ), а их стремительная адаптация содействует освобождению восприятия от на биологическом уровне малозначительной инфы (к примеру, контакт меж кожей и одежкой). Возбуждение медлительно адаптирующихся рецепторов не достаточно находится в зависимости от скорости конфигурации стимула и сохраняется при его продолжительном действии (тонический ответ), потому, к примеру, неспешная адаптация проприоцепторов позволяет человеку получать подходящую ему для сохранения позы информацию в течение всего нужного времени.
Есть сенсорные нейроны, генерирующие потенциалы деяния спонтанно, т. е. при отсутствии раздражения (к примеру, сенсорные нейроны вестибулярной системы), такая активность именуется фоновой. Частота нервных импульсов в этих нейронах может возрастать либо уменьшаться зависимо от интенсивности действующего на вторичные сенсоры стимула, не считая того, она может определяться направлением, в каком отклоняются чувствительные волоски механорецепторов. К примеру, отклонение волосков вторичных механорецепторов в одну сторону сопровождается увеличением фоновой активности сенсорного нейрона, которому они принадлежат, а в обратную сторону — снижением его фоновой активности. Обозначенный метод рецепции позволяет получать информацию и об интенсивности стимула, и о направлении, в каком он действует.
Рецептивные поля органов эмоций живых систем
Рецептивным полем именуется область, занимаемая совокупой всех рецепторов, стимуляция которых приводит к возбуждению сенсорного нейрона (рис. 17.1). Наибольшая величина рецептивного поля первичного сенсорного нейрона определяется местом, которое занимают все ветки его периферического отростка, а число рецепторов, имеющихся в этом пространстве, показывает на плотность иннервации. Высочайшая плотность иннервации смешивается, обычно, с малыми размерами рецептивных полей и, соответственно, высочайшим пространственным разрешением, позволяющим различать стимулы, действующие на примыкающие рецептивные поля. Малые рецептивные поля типичны, к примеру, для центральной ямки сетчатки и для пальцев рук, где плотность рецепторов существенно выше, чем на периферии сетчатки либо в коже спины, для которых свойственны большая величина рецептивных полей и наименьшее пространственное разрешение. Рецептивные поля примыкающих сенсорных нейронов могут отчасти перекрывать друг дружку, потому информация о действующих на их стимулах передается не по одному, а по нескольким параллельным аксонам, что увеличивает надежность ее передачи.
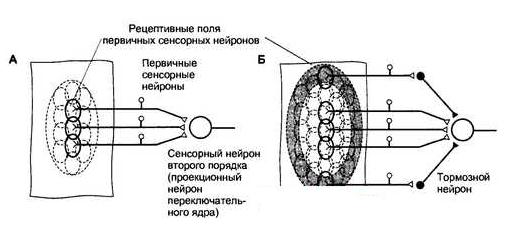
Рис. 12.2 Рецептивные поля первичных сенсорных нейронов и сенсорных нейронов второго порядка. Рецептивные поля первичных сенсорных нейронов ограничены областью их чувствительных окончаний. Рецептивное поле переключательного нейрона появляется из суммы рецептивных полей конвергирующих к нему первичных сенсорных нейронов. Раздражение центральной либо периферической области рецептивного поля сенсорного нейрона второго и последующих порядков сопровождается обратным эффектом. Как видно на схеме, раздражение центра рецептивного поля вызовет возбуждение проекционного нейрона, а раздражение периферической области — торможение при помощи интернейронов переключательного ядра (латеральное торможение). В итоге контраста, создаваемого меж центром и периферией рецептивного поля, выделяется информация для передачи на последующий иерархический уровень.
Величина рецептивных полей сенсорных нейронов второго и последующих порядков больше, чем у первичных сенсорных нейронов, так как центральные нейроны получают информацию от нескольких конвергирующих к ним нейронов предыдущего уровня. От центра рецептивного поля информация передается конкретно к сенсорным нейронам последующего порядка, а от периферии — к тормозным интернейронам переключательного ядра, потому центр и периферия рецептивного поля являются реципрокными по отношению друг к другу. В итоге сигналы от центра рецептивного поля беспрепятственно добиваются последующего иерархического уровня сенсорной системы, тогда как сигналы, поступающие от периферии рецептивного поля, тормозятся (в другом варианте организации рецептивного поля легче пропускаются сигналы от периферии, а не от центра). Такая многофункциональная организация рецептивных полей обеспечивает выделение более важных сигналов, просто различаемых на контрастном с ними фоне.
Сенсорные нейроны низшего иерархического уровня способны передавать электронные сигналы сразу нескольким нейронам последующего уровня благодаря дивергенции к ним собственных аксонов. Это увеличивает надежность передачи инфы от 1-го иерархического уровня к другому и позволяет сохранять чувствительность сенсорной системы при утрате отдельных нейронов. Возбуждение нейронов более высочайшего иерархического уровня определяется не только лишь действием на их нейронов предшествующего уровня, да и механизмом латерального торможения, имеющегося в переключательных ядрах (рис. 12.3).
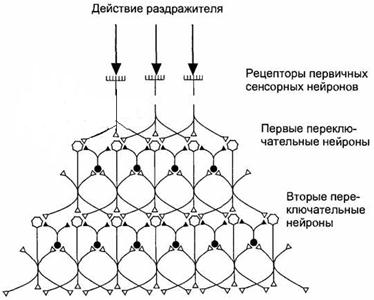
Рис. 12.3 Схема латерального торможения в сенсорных системах.
Степень возбуждения переключательных нейронов определяется интенсивностью раздражения центра либо периферии его рецептивного поля. В итоге возбуждение будет передаваться от одной части рецептивного поля на фоне торможения либо прерывания сигналов от другой его части, что делает эффект контрастирования, содействующий различению действующих стимулов.
Латеральное торможение происходит вследствие активации тормозных интернейронов переключательного ядра коллатералями возбуждающих нейронов. Чем посильнее возбужден релейный сенсорный нейрон, тем больше он активирует тормозные нейроны, которые подавляют активность примыкающих релейных нейронов. Релейные нейроны, передающие сигналы от центра рецептивного поля, посильнее других увеличивают активность тормозных нейронов, а их воздействие на примыкающие релейные нейроны делает еще огромным контраст меж возбужденными и тормозимыми нейронами, выделяя линию передачи избранных сигналов.
Механизмы реализации отрицательной и положительной оборотных связей
Высшие уровни сенсорной системы регулируют переработку инфы в низших по отношению к ним переключательных ядрах средством нисходящего торможения либо усиления передаваемых сигналов. Нисходящее торможение (рис.12.3) происходит вследствие активации тормозных нейронов переключательного ядра, деятельность которых определяют нисходящие пути, начинающиеся на более высочайшем иерархическом уровне сенсорной системы. В итоге нисходящего торможения увеличивается порог афферентной синаптической передачи в переключательном ядре низшего уровня. Нисходящее торможение представляет собой регуляцию сенсорного восприятия по механизму отрицательной оборотной связи. Тормозные интернейроны переключательных ядер уменьшают частоту передаваемых на последующий уровень сигналов, в связи с чем, сенсорное чувство ослабляется.
Нисходящее усиление происходит при помощи возбуждающих интернейронов переключательного ядра, которые активизируются аксонами нейронов более высочайшего иерархического уровня. Вследствие этого в управляемом по механизму положительной оборотной связи переключательном ядре снижается порог синаптической передачи, что делает вероятным прохождение относительно слабеньких сигналов на последующий иерархический уровень. Одновременное внедрение устройств отрицательной и положительной оборотной связи позволяет освобождаться от лишней инфы, подавлять «шум», т. е. нейронную активность, не содержащую физиологически важных сообщений, и сразу выделять и усиливать сигналы, на которых будет сосредоточено внимание.
Большая часть раздражителей, воспринимаемых сенсорными системами, оказывает на их общее воздействие, так как сразу возбуждает различные типырецепторов. К примеру, прикасающийся к ограниченному участку кожи предмет может быть гладким либо шершавым, теплым либо прохладным, сухим либо мокроватым, а его давление на кожу может усиливаться либо слабеть, он может передвигаться в том либо ином направлении.
Зрительные чувства представляют собой сочетание определенной формы воспринимаемого предмета, его движения и композиции разных цветов, заполняющих ту либо иную деталь формы. Сенсорные системы организованы так, что информация о каждом компоненте всеохватывающего раздражителя передается сразу по нескольким параллельным путям, любой из которых относится к определенной субмодальности, как, к примеру, форма, движение либо цвет наблюдаемого зрительного объекта. В каждом из таких путей независимо друг от друга перерабатываются сигналы, несущие информацию о различных качествах подействовавшего на сенсоры всеохватывающего раздражителя. При всем этом различные каналы, применяемые для передачи сенсорной инфы, не дублируют друг дружку, в чем проявляется принцип многоканальности, присущий всем сенсорным системам. Высочайшая упорядоченность нейронных переключений на каждом иерархическом уровне обеспечивает поступление инфы от каждого рецептивного поля к подходящим конкретно ему кортикальным колонкам проекционной коры. Нужное для целостного восприятия раздражителя объединение инфы, относящейся к различным субмодальностям, происходит в сенсорных регионах коры.
Личное сенсорное восприятие, абсолютный порог чувства. Закон Вебера—Фехнера.
Любая сенсорная система принимает действие адекватного стимула в ограниченном спектре значений его силы. Меньший по интенсивности стимул, способный вызвать чувство, именуется абсолютным порогом чувства. Его величина устанавливается опытным методом, она не схожа у различных людей и может изменяться у 1-го и такого же человека зависимо от многофункционального состояния, различаясь, к примеру, при хорошей работоспособности и утомлении. Величина абсолютного порога чувства выше абсолютного порога первичных сенсорных нейронов, потому что возбуждение отдельных чувствительных нейронов либо их маленький группы не непременно приводит к возбуждению высших уровней сенсорной системы вследствие тормозных процессов на прошлых иерархических уровнях. Потому не каждый стимул, возбуждающий сенсоры первичного сенсорного нейрона, лично чувствуется и осознается.
Величина, на которую один стимул надпорогового спектра должен отличаться от другого, чтоб их разницу можно было лично различить, получила заглавие дифференциального порога либо порога различения. Ощущаемый прирост интенсивности раздражения (дифференциальный порог) должен превосходить ранее действовавший стимул на определенную и постоянную величину.
Закон Вебера справедлив для различных модальностей, но только в границах раздражения средней силы, а при слабеньких и сильных стимулах способность к различению силы стимулов у человека понижается. Предложена также ступенчатая шкала интенсивности чувств, в какой за ноль принимается величина абсолютного порога, а зависимость меж интенсивностью стимула и чувством выражается как:
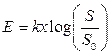 , (12.1)
, (12.1)
где Е — интенсивность чувства, к — неизменная величина, S — интенсивность стимула, 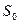 — абсолютный порог.
— абсолютный порог.
Эта зависимость, получившая заглавие закона Вебера—Фехнера, указывает, что линейное повышение интенсивности чувства отражает логарифмический рост интенсивности стимула. Закон Вебера—Фехнера охарактеризовывает личную способность человека различать действующие стимулы (шкала различения), но не оценивает саму интенсивность чувств, так как этот закон основан на допущении равенства всех ее приростов, как к слабеньким, так и к сильным стимулам.
Личная оценка интенсивности раздражителя (высококачественное различие) растет при увеличении различия меж пороговой и действующей силой стимула (количественное различие). Но зависимость меж силой стимула и чувством не схожа при разной интенсивности стимулов, а поэтому имеет не линейный, а степенной нрав. Для оценки интенсивности чувств всего спектра употребляется шкала Стивенса, устанавливающая зависимость чувств от силы стимула в виде степенной функции:
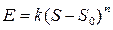 , (12.2)
, (12.2)
где Е — интенсивность чувства, S — действующая сила стимула, 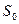 — абсолютный порог, к — константа шкалы, n — показатель степени, который находится в зависимости от сенсорной модальности (к примеру, для восприятия светового чувства он составляет 0,33, громкости звука — 0,6, а для проприоцептивных чувств — 1,7).
— абсолютный порог, к — константа шкалы, n — показатель степени, который находится в зависимости от сенсорной модальности (к примеру, для восприятия светового чувства он составляет 0,33, громкости звука — 0,6, а для проприоцептивных чувств — 1,7).
Пространственные свойства действующих стимулов, нужные для их различения, зависят от специфичных особенностей каждой сенсорной системы и величины рецептивных полей. Прикосновение к коже дистальной фаланги пальца руки 2-ух ножек циркуля с расстоянием меж ними 2 мм чувствуется раздельно, но чтоб почувствовать раздельное прикосновение к коже спины, ножки циркуля нужно раздвинуть до 60 мм. Пространственное восприятие этих тактильных стимулов находится в зависимости от размеров соответственных рецептивных полей: раздельное чувство может быть только при условии раздражения каждой ножкой циркуля независящего рецептивного поля. Только тогда информация о каждом стимуле будет перерабатываться раздельно на каждом уровне организации сенсорной системы, включая проекционную область коры. Подобная ситуация имеет место при восприятии 2-ух точек зрительного поля: они не соединяются в одну, если отражаемые ими световые лучи попадут на различные рецептивные поля сетчатки. Имеет значение и степень контраста меж действующим стимулом и его фоном: отлично контрастируемые объекты (к примеру, темное на белоснежном) различаются легче, чем не достаточно контрастируемые (темное на сероватом).
Рецептивные поля различающихся своими сенсорами сенсорных нейронов перекрываются, потому при действии на кожу комплекса стимулов сразу возбуждаются различные виды рецепторов, что позволяет чувствовать все динамические и статические характеристики такового комплекса. Обработка и анализ инфы сигналов от разных рецепторов происходит на высших уровнях сенсорной системы, формирующих всеохватывающее восприятие действующих на кожа стимулов. Плотность механорецепторов в различных участках кожи не схожа, чем определяются различные характеристики пространственного дифференциального порога, т. е. меньшего расстояния меж 2-мя точками, раздражение каждой из которых чувствуется раздельно.